Пластиды - органоиды, специфичные для клеток растений (они имеются в клетках всех растений, за исключением большинства бактерий, грибов и некоторых водорослей).
В клетках высших растений находится обычно от 10 до 200 пластид размером 3-10мкм, чаще всего имеющих форму двояковыпуклой линзы. У водорослей зеленые пластиды, называемые хроматофорами, очень разнообразны по форме и величине. Они могут иметь звездчатую, лентовидную, сетчатую и другие формы.
Различают 3 вида пластид:
- Бесцветные пластиды - лейкопласты ;
- окрашенные - хлоропласты (зеленого цвета);
- окрашенные - хромопласты (желтого, красного и других цветов).
Эти виды пластид до известной степени способны превращаться друг в друга - лейкопласты при накоплении хлорофилла переходят в хлоропласты, а последние при появлении красных, бурых и других пигментов - в хромопласты.
Строение и функции хлоропластов
Хлоропласты - зеленые пластиды, содержащие зеленый пигмент - хлорофилл.
Основная функция хлоропласт - фотосинтез.
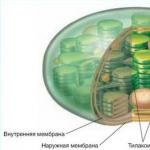
В хлоропластах есть свои рибосомы, ДНК, РНК, включения жира, зерна крахмала. Снаружи хлоропласта покрыты двумя белково-липидными мембранами, а в их полужидкую строму (основное вещество) погружены мелкие тельца - граны и мембранные каналы.

Граны (размером около 1мкм) - пакеты круглых плоских мешочков (тилакоидов), сложенных подобно столбику монет. Располагаются они перпендикулярно поверхности хлоропласта. Тилакоиды соседних гран соединены между собой мембранными каналами, образуя единую систему. Число гран в хлоропластах различно. Например, в клетках шпината каждый хлоропласт содержит 40-60 гран.
Хлоропласты внутри клетки могут двигаться пассивно, увлекаемые током цитоплазмы, либо активно перемещаться с места на место.
- Если свет очень интенсивен, они поворачиваются ребром к ярким лучам солнца и выстраиваются вдоль стенок, параллельных свету.
- При слабом освещении, хлоропласты перемещаются на стенки клетки, обращенные к свету, и поворачиваются к нему своей большой поверхностью.
- При средней освещенности они занимают среднее положение.
Этим достигаются наиболее благоприятные для процесса фотосинтеза условия освещения.
Хлорофилл
В гранах пластид растительной клетки содержится хлорофилл, упакованный с белковыми и фосфолипидными молекулами так, чтобы обеспечить способность улавливать световую энергию.
Молекула хлорофилла очень сходна с молекулой гемоглобина и отличается главным образом тем, что расположенный в центре молекулы гемоглобина атом железа заменен в хлорофилле на атом магния.

В природе встречается четыре типа хлорофилла: a, b, c, d.
Хлорофиллы a и b содержат высшие растения и зеленые водоросли, диатомовые водоросли содержат a и c, красные - a и d.
Лучше других изучены хлорофиллы a и b (их впервые разделил русский ученый М.С.Цвет в начале XXв.). Кроме них существуют четыре вида бактериохлорофиллов - зеленых пигментов пурпурных и зеленых бактерий: a, b, c, d.
Большинство фотосинтезирующих бактерий содержат бактериохлорофилл a, некоторые - бактериохлорофилл b, зеленые бактерии - c и d.
Хлорофилл обладает способностью очень эффективно поглощать солнечную энергию и передавать ее другим молекулам, что является его главной функцией. Благодаря этой способности хлорофилл - единственная структура на Земле, которая обеспечивает процесс фотосинтеза.
Главная функция хлорофилла в растениях - поглощение энергии света и передача ее другим клеткам.
Пластидам, так же, как и митохондриям, свойственна до некоторой степени автономность внутри клетки. Они размножаются путем деления.
Наряду с фотосинтезом, в пластидах происходит процесс биосинтеза белка. Благодаря содержанию ДНК пластиды играют определенную роль в передаче признаков по наследству (цитоплазматическая наследственность).
Строение и функции хромопластов
Хромопласты относятся к одному из трех видов пластид высших растений. Это небольших размеров, внутриклеточные органеллы.
Хромопласты имеют различный окрас: желтый, красный, коричневый. Они придают характерный цвет созревшим плодам, цветкам, осенней листве. Это необходимо для привлечения насекомых-опылителей и животных, которые питаются плодами и разносят семена на дальние расстояния.

Структура хромопласта похожа на другие пластиды. Их двух оболочек внутренняя развита слабо, иногда вовсе отсутствует. В ограниченном пространстве расположена белковая строма, ДНК и пигментные вещества (каротиноиды).
Каротиноиды – это жирорастворимые пигменты, которые накапливаются в виде кристаллов.
Форма хромопластов очень разнообразна: овальная, многоугольная, игольчатая, серповидная.
Роль хромопластов в жизни растительной клетки до конца не выяснена. Исследователи предполагают, что пигментные вещества играют важную роль в окислительно-восстановительных процессах, необходимы для размножения и физиологичного развития клетки.
Строение и функции лейкопластов
Лейкопласты - это органоиды клетки, в которых накапливаются питательные вещества. Органеллы имеют две оболочки: гладкую наружную и внутреннюю с несколькими выступами.
Лейкопласты на свету превращаются в хлоропласты (к примеру зеленые клубни картофеля), в обычном состоянии они бесцветны.
Форма лейкопластов шаровидная, правильная. Они находятся в запасающей ткани растений, которая заполняет мягкие части: сердцевину стебля, корня, луковиц, листьев.

Функции лейкопластов зависят от их вида (в зависимости от накапливаемого питательного вещества).
Разновидности лейкопластов:
- Амилопласты накапливают крахмал, встречаются во всех растениях, так как углеводы основной продукт питания растительной клетки. Некоторые лейкопласты полностью наполнены крахмалом, их называют крахмальными зернами.
- Элайопласты продуцируют и запасают жиры.
- Протеинопласты содержат белковые вещества.
Лейкопласты также служат ферментной субстанцией. Под действием ферментов быстрее протекают химические реакции. А в неблагоприятный жизненный период, когда процессы фотосинтеза не осуществляются, они расщепляют полисахариды до простых углеводов, которые необходимы растениям для выживания.
В лейкопластах не может происходить фотосинтез, потому что они не содержат гран и пигментов.
Луковицы растений, в которых содержится много лейкопластов, могут переносить длительные периоды засухи, низкую температуру, жару. Это связано с большими запасами воды и питательных веществ в органеллах.
Предшественниками всех пластид является пропластиды, небольшие органоиды. Допускают, что лейко — и хлоропласты способны трансформироваться в другие виды. В конечном итоге после выполнения своих функций хлоропласты и лейкопласты становятся хромопластами — это последняя стадия развития пластид.
Важно знать! Одновременно в клетке растения может находиться только один вид пластид.
Сводная таблица строения и функций пластид
| Свойства | Хлоропласты | Хромопласты | Лейкопласты |
|---|---|---|---|
| Строение | Двухмембранная органелла, с гранами и мембранными канальцами | Органелла с не развитой внутренней мембранной системой | Мелкие органеллы, находятся в частях растения, скрытых от света |
| Окрас | Зеленые | Разноцветные | Бесцветные |
| Пигмент | Хлорофилл | Каротиноид | Отсутствует |
| Форма | Округлая | Многоугольная | Шаровидная |
| Функции | Фотосинтез | Привлечение потенциальных распространителей растений | Запас питательных веществ |
| Заменимость | Переходят в хромопласты | Не изменяются, это последняя стадия развития пластид | Превращаются в хлоропласты и хромопласты |
Это бесцветные или окрашенные тельца в протоплазме растительных клеток, представляющие собой сложную систему внутренних мембран (мембранные органеллы) и выполняющие различные функции. Бесцветные пластиды называют лейкопластами , различно окрашенные (желтого, оранжевого или красного цвета) - хромопластами , зеленые - хлоропластами . В клетке высших растений содержится около 40 хлоропластов в которых происходит фотосинтез . Они, как уже было сказано, способны к автономному размножению, не зависящему от деления клетки. Размеры и форма митохондрий и хлоропластов, наличие в их матриксе кольцевых двухцепочных ДНК и собственных рибосом делают эти органеллы похожими на бактериальные клетки. Существует теория симбиотического происхождения эукариотической клетки , согласно которой предки современных митохондрий и хлоропластов были когда-то самостоятельными прокариотическими организмами.
Пластиды характерны только для растений. Они не найдены у грибов и у большинства животных, исключая некоторых фотосинтезирующих простейших.
Предшественниками пластид являются пропластиды , мелкие, обычно бесцветные образования, находящиеся в делящихся клетках корней и побегов . Если развитие пропластид в более дифференцированные структуры задерживается из-за отсутствия света, в них может появиться одно или несколько проламеллярных телец (скопления трубчатых мембран). Такие бесцветные пластиды называются этиопластами . Этиопласты превращаются в хлоропласты на свету, а из мембран проламеллярных телец формируются тилакоиды . В зависимости от окраски, связанной с наличием или отсутствием тех или иных пигментов, различают три основных типа пластид (см. выше) - хлоропласты , хромопласты и лейкопласты . Обычно в клетке встречаются пластиды только одного типа. Однако установлено, что одни типы пластид могут переходить в другие.
Пластиды - относительно крупные образования клетки. Самые большие из них - хлоропласты - достигают у высших растений 4-10 мкм длины и хорошо различимы в световой микроскоп. Форма окрашенных пластид чаще всего линзовидная или эллиптическая. В клетках встречаются, как правило, несколько десятков пластид, но у водорослей, где пластиды нередко крупны и разнообразны по форме, число их иногда невелико (1-5). Такие пластиды называются хроматофорами . Лейкопласты и хромопласты могут иметь различную форму.
Основная функция хлоропластов - фотосинтез. Центральная роль в этом процессе принадлежит хлорофиллу , точнее - нескольким его модификациям. Световые реакции фотосинтеза осуществляются преимущественно в гранах , темновые - в строме
эукариотических растений и некоторых фотосинтезирующих простейших. Покрыты двойной мембраной и имеют в своём составе множество копий кольцевой ДНК . Совокупность пластид клетки образует пластидом. По окраске и выполняемой функции выделяют три основных типа пластид:- Лейкопласты - неокрашенные пластиды, как правило выполняют запасающую функцию. В лейкопластах клубней картофеля накапливается крахмал. Лейкопласты высших растений могут превращаться в хлоропласты или хромопласты.
- Хромопласты - пластиды, окрашенные в жёлтый, красный или оранжевый цвет. Окраска хромопластов связана с накоплением в них каротиноидов . Хромопласты определяют окраску осенних листьев, лепестков цветов, корнеплодов, созревших плодов.
- Хлоропласты - пластиды, несущие фотосинтезирующие пигменты - хлорофиллы . Имеют зелёную окраску у высших растений, харовых и зелёных водорослей. Набор пигментов, участвующих в фотосинтезе (и, соответственно, определяющих окраску хлоропласта) различен у представителей разных таксономических отделов. Хлоропласты имеют сложную внутреннюю структуру
Wikimedia Foundation . 2010 .
Синонимы :Смотреть что такое "Пластида" в других словарях:
ПЛАСТИДА, разновидность ОРГАНОИДОВ, находящихся в КЛЕТКАХ растений и зеленых водорослей. Пластиды имеют двойную мембрану и содержат ДНК. Среди примеров пластид можно назвать ХЛОРОПЛАСТЫ и лейкопласты … Научно-технический энциклопедический словарь
Сущ., кол во синонимов: 5 амилопласт (2) органоид (5) хлоропласт (2) … Словарь синонимов
пластида - Органоид эукариотической растительной клетки или автономная самоудваивающаяся частица растительного организма Тематики биотехнологии EN plastid … Справочник технического переводчика
Plastid пластида. Самовоспроизводящаяся клеточная органелла эукариотических растительных клеток; различают 3 основные группы П. хлоропласты (зеленые), лейкопласты (бесцветные) и хромопласты (желтые и красные); многие П. обладают собственным… … Молекулярная биология и генетика. Толковый словарь.
Ж. Окрашенное или бесцветное живое тельце, находящееся в протоплазме клеток растений. Толковый словарь Ефремовой. Т. Ф. Ефремова. 2000 … Современный толковый словарь русского языка Ефремовой
пластида - и, ж. Особливе живе тільце в протоплазмі рослинних клітин … Український тлумачний словник
, бурые , жёлто-зелёные , диатомовые водоросли) мембран считается результатом двух- и трёхкратного эндосимбиоза соответственно.
Общие черты строения пластид высших растений
Типичные пластиды высших растений окружены оболочкой из двух мембран - внешней и внутренней. Внутренняя и внешняя мембраны пластид бедны фосфолипидами и обогащены галактолипидами . Внешняя мембрана не имеет складок, никогда не сливается с внутренней мембраной и содержит поровый белок, обеспечивающий свободный транспорт воды, ионов и метаболитов с массой до 10 кДа. Внешняя мембрана имеет зоны тесного контакта с внутренней мембраной; предполагается, что в этих участках осуществляется транспорт белков из цитоплазмы в начале пластид. Внутренняя мембрана проницаема для небольших незаряженных молекул и для недиссоциированных низкомолекулярных монокарбоновых кислот, для более крупных и заряженных метаболитов в мембране локализованы белковые переносчики. Строма - внутреннее содержимое пластид - представляет собой гидрофильный матрикс, содержащий неорганические ионы, водорастворимые органические метаболиты, геном пластид (несколько копий кольцевой ДНК), рибосомы прокариотического типа, ферменты матричного синтеза и другие ферментативные системы. Эндомембранная система пластид развивается в результате отшнуровки везикул от внутренней мембраны и их упорядочивания. Степень развития эндомембранной системы зависит от типа пластид. Наибольшего развития эндомембранная система достигает в хлоропластах, где она является местом протекания световых реакций фотосинтеза и представлена свободными тилакоидами стромы и тилакоидами, собранными в стопки - граны . Внутреннее пространство эндомембран называется люмен. Люмен тилакоидов, также как и строма, содержит ряд водорастворимых белков.
Геном и белоксинтезирующая система пластид высших растений
Одним из доказательств происхождения пластид от древних цианобактерий служит схожесть их геномов, хотя пластидный геном (пластом) значительно меньше. Пластом высших растений представлен многокопийной кольцевой двуцепочечной ДНК (плДНК) размером от 75 до 290 тыс. п. н. В большинстве пластидных геномов присутствуют два инвертированных повтора (IR A и IR B), разделяющих молекулу ДНК на две уникальные области: большую (LSR) и малую (SSR). В инвертированных повторах содержатся гены всех четырёх рРНК (4,5S, 5S, 16S и 23S), входящих в состав пластидных рибосом, а также гены некоторых тРНК . Голосеменные и растения семейства Бобовые не содержат инвертированных повторов. Многие пластидные гены организованы в опероны - группы генов, считывающихся с общего промотора . Некоторые пластидные гены имеют экзон-интронную структуру. В пластидах кодируются гены, обслуживающие процессы транскрипции и трансляции (гены «домашнего хозяйства»), а также некоторые гены, обеспечивающие выполнение функций пластид в клетке, прежде всего фотосинтез .
Транскрипцию в пластидах обеспечивают РНК-полимеразы двух типов:
- Мультисубъединичная пластидная РНК-полимераза бактериального типа состоит из двух α-субъединиц и по одной β, β", β" (все эти субъединицы кодируются в пластидном геноме). Однако для её активации необходимо присутствие σ-субъединицы, которая кодируется в ядре растительной клетке и импортируется в пластиды при освещении. Таким образом пластидная РНК-полимераза активна только на свету. Пластидная РНК-полимераза может обеспечивать транскрипцию с генов с эубактериальными промоторами (большинство генов фотосинтетических белков), а также с генов, имеющих универсальные промоторы.
- Мономерная РНК-полимераза фагового типа кодируется в ядре и белок имеет специальную сигнальную последовательность обеспечивающую импорт в пластиды. Обеспечивает транскрипцию генов «домашнего хозяйства» (в частности гены rif-оперона, который содержит гены пластидной РНК-полимеразы).
Процесс созревания транскриптов пластид имеет свои особенности. В частности пластидные интроны способны к автосплайсингу , то есть вырезания интронов происходит автокаталитически. Кроме того в пластидах происходит редактирование РНК - химическая модификация оснований РНК, приводящая к изменению закодированной информации (наиболее часто происходит замена цитидина на уридин) . Большинство зрелых мРНК пластид содержат в 3"-некодирующей области шпильку, защищающую её от рибонуклеаз .
- Хлоропласты - зелёные пластиды, основной функцией которых является фотосинтез. Хлоропласты как правило имеют элипсовидную форму и длину от 5 до 8 мкм. Количество хлоропластов в клетке различно: в клетке хлоренхимы листа Arabidopsis содержится около 120 хлоропластов, в губчатой хлоренхиме листа клещевины их около 20, клетка нитчатой морской водоросли Spirogyra содержит единственный лентовидный хлоропласт. Хлоропласты имеют хорошо развитую эндомембранную систему, в которой выделяют тилакоиды стромы и стопки тилакоидов - граны. Зелёная окраска хлоропластов обусловлена высоким содержанием основного пигмента фотосинтеза - хлорофилла . Помимо хлорофилла хлоропласты содержат различные каротиноиды. Набор пигментов, участвующих в фотосинтезе (и, соответственно окраска) различен у представителей разных таксонов.
- Хромопласты - пластиды, окрашенные в жёлтый, красный или оранжевый цвет. Хромопласты могут развиваться из пропластид или повторно дифференцироваться из хлоропластов; также хромопласты могут редифференцироваться в хлоропласты. Окраска хромопластов связана с накоплением в них каротиноидов . Хромопласты определяют окраску осенних листьев, лепестков некоторых цветов (лютики, бархатцы), корнеплодов (морковь), созревших плодов (томат).
Напишите отзыв о статье "Пластиды"
Ссылки
|
||||||||||||||||||||||||||||

|
||||||||||||||||||||||
Отрывок, характеризующий Пластиды
Дрон, не отвечая, вздохнул.– Если прикажете, они уйдут, – сказал он.
– Нет, нет, я пойду к ним, – сказала княжна Марья
Несмотря на отговариванье Дуняши и няни, княжна Марья вышла на крыльцо. Дрон, Дуняша, няня и Михаил Иваныч шли за нею. «Они, вероятно, думают, что я предлагаю им хлеб с тем, чтобы они остались на своих местах, и сама уеду, бросив их на произвол французов, – думала княжна Марья. – Я им буду обещать месячину в подмосковной, квартиры; я уверена, что Andre еще больше бы сделав на моем месте», – думала она, подходя в сумерках к толпе, стоявшей на выгоне у амбара.
Толпа, скучиваясь, зашевелилась, и быстро снялись шляпы. Княжна Марья, опустив глаза и путаясь ногами в платье, близко подошла к ним. Столько разнообразных старых и молодых глаз было устремлено на нее и столько было разных лиц, что княжна Марья не видала ни одного лица и, чувствуя необходимость говорить вдруг со всеми, не знала, как быть. Но опять сознание того, что она – представительница отца и брата, придало ей силы, и она смело начала свою речь.
– Я очень рада, что вы пришли, – начала княжна Марья, не поднимая глаз и чувствуя, как быстро и сильно билось ее сердце. – Мне Дронушка сказал, что вас разорила война. Это наше общее горе, и я ничего не пожалею, чтобы помочь вам. Я сама еду, потому что уже опасно здесь и неприятель близко… потому что… Я вам отдаю все, мои друзья, и прошу вас взять все, весь хлеб наш, чтобы у вас не было нужды. А ежели вам сказали, что я отдаю вам хлеб с тем, чтобы вы остались здесь, то это неправда. Я, напротив, прошу вас уезжать со всем вашим имуществом в нашу подмосковную, и там я беру на себя и обещаю вам, что вы не будете нуждаться. Вам дадут и домы и хлеба. – Княжна остановилась. В толпе только слышались вздохи.
– Я не от себя делаю это, – продолжала княжна, – я это делаю именем покойного отца, который был вам хорошим барином, и за брата, и его сына.
Она опять остановилась. Никто не прерывал ее молчания.
– Горе наше общее, и будем делить всё пополам. Все, что мое, то ваше, – сказала она, оглядывая лица, стоявшие перед нею.
Все глаза смотрели на нее с одинаковым выражением, значения которого она не могла понять. Было ли это любопытство, преданность, благодарность, или испуг и недоверие, но выражение на всех лицах было одинаковое.
– Много довольны вашей милостью, только нам брать господский хлеб не приходится, – сказал голос сзади.
– Да отчего же? – сказала княжна.
Никто не ответил, и княжна Марья, оглядываясь по толпе, замечала, что теперь все глаза, с которыми она встречалась, тотчас же опускались.
– Отчего же вы не хотите? – спросила она опять.
Никто не отвечал.
Княжне Марье становилось тяжело от этого молчанья; она старалась уловить чей нибудь взгляд.
– Отчего вы не говорите? – обратилась княжна к старому старику, который, облокотившись на палку, стоял перед ней. – Скажи, ежели ты думаешь, что еще что нибудь нужно. Я все сделаю, – сказала она, уловив его взгляд. Но он, как бы рассердившись за это, опустил совсем голову и проговорил:
– Чего соглашаться то, не нужно нам хлеба.
– Что ж, нам все бросить то? Не согласны. Не согласны… Нет нашего согласия. Мы тебя жалеем, а нашего согласия нет. Поезжай сама, одна… – раздалось в толпе с разных сторон. И опять на всех лицах этой толпы показалось одно и то же выражение, и теперь это было уже наверное не выражение любопытства и благодарности, а выражение озлобленной решительности.
– Да вы не поняли, верно, – с грустной улыбкой сказала княжна Марья. – Отчего вы не хотите ехать? Я обещаю поселить вас, кормить. А здесь неприятель разорит вас…
Но голос ее заглушали голоса толпы.
– Нет нашего согласия, пускай разоряет! Не берем твоего хлеба, нет согласия нашего!
Княжна Марья старалась уловить опять чей нибудь взгляд из толпы, но ни один взгляд не был устремлен на нее; глаза, очевидно, избегали ее. Ей стало странно и неловко.
– Вишь, научила ловко, за ней в крепость иди! Дома разори да в кабалу и ступай. Как же! Я хлеб, мол, отдам! – слышались голоса в толпе.
Княжна Марья, опустив голову, вышла из круга и пошла в дом. Повторив Дрону приказание о том, чтобы завтра были лошади для отъезда, она ушла в свою комнату и осталась одна с своими мыслями.
Долго эту ночь княжна Марья сидела у открытого окна в своей комнате, прислушиваясь к звукам говора мужиков, доносившегося с деревни, но она не думала о них. Она чувствовала, что, сколько бы она ни думала о них, она не могла бы понять их. Она думала все об одном – о своем горе, которое теперь, после перерыва, произведенного заботами о настоящем, уже сделалось для нее прошедшим. Она теперь уже могла вспоминать, могла плакать и могла молиться. С заходом солнца ветер затих. Ночь была тихая и свежая. В двенадцатом часу голоса стали затихать, пропел петух, из за лип стала выходить полная луна, поднялся свежий, белый туман роса, и над деревней и над домом воцарилась тишина.
Одна за другой представлялись ей картины близкого прошедшего – болезни и последних минут отца. И с грустной радостью она теперь останавливалась на этих образах, отгоняя от себя с ужасом только одно последнее представление его смерти, которое – она чувствовала – она была не в силах созерцать даже в своем воображении в этот тихий и таинственный час ночи. И картины эти представлялись ей с такой ясностью и с такими подробностями, что они казались ей то действительностью, то прошедшим, то будущим.
То ей живо представлялась та минута, когда с ним сделался удар и его из сада в Лысых Горах волокли под руки и он бормотал что то бессильным языком, дергал седыми бровями и беспокойно и робко смотрел на нее.
«Он и тогда хотел сказать мне то, что он сказал мне в день своей смерти, – думала она. – Он всегда думал то, что он сказал мне». И вот ей со всеми подробностями вспомнилась та ночь в Лысых Горах накануне сделавшегося с ним удара, когда княжна Марья, предчувствуя беду, против его воли осталась с ним. Она не спала и ночью на цыпочках сошла вниз и, подойдя к двери в цветочную, в которой в эту ночь ночевал ее отец, прислушалась к его голосу. Он измученным, усталым голосом говорил что то с Тихоном. Ему, видно, хотелось поговорить. «И отчего он не позвал меня? Отчего он не позволил быть мне тут на месте Тихона? – думала тогда и теперь княжна Марья. – Уж он не выскажет никогда никому теперь всего того, что было в его душе. Уж никогда не вернется для него и для меня эта минута, когда бы он говорил все, что ему хотелось высказать, а я, а не Тихон, слушала бы и понимала его. Отчего я не вошла тогда в комнату? – думала она. – Может быть, он тогда же бы сказал мне то, что он сказал в день смерти. Он и тогда в разговоре с Тихоном два раза спросил про меня. Ему хотелось меня видеть, а я стояла тут, за дверью. Ему было грустно, тяжело говорить с Тихоном, который не понимал его. Помню, как он заговорил с ним про Лизу, как живую, – он забыл, что она умерла, и Тихон напомнил ему, что ее уже нет, и он закричал: „Дурак“. Ему тяжело было. Я слышала из за двери, как он, кряхтя, лег на кровать и громко прокричал: „Бог мой!Отчего я не взошла тогда? Что ж бы он сделал мне? Что бы я потеряла? А может быть, тогда же он утешился бы, он сказал бы мне это слово“. И княжна Марья вслух произнесла то ласковое слово, которое он сказал ей в день смерти. «Ду ше нь ка! – повторила княжна Марья это слово и зарыдала облегчающими душу слезами. Она видела теперь перед собою его лицо. И не то лицо, которое она знала с тех пор, как себя помнила, и которое она всегда видела издалека; а то лицо – робкое и слабое, которое она в последний день, пригибаясь к его рту, чтобы слышать то, что он говорил, в первый раз рассмотрела вблизи со всеми его морщинами и подробностями.
«Душенька», – повторила она.
«Что он думал, когда сказал это слово? Что он думает теперь? – вдруг пришел ей вопрос, и в ответ на это она увидала его перед собой с тем выражением лица, которое у него было в гробу на обвязанном белым платком лице. И тот ужас, который охватил ее тогда, когда она прикоснулась к нему и убедилась, что это не только не был он, но что то таинственное и отталкивающее, охватил ее и теперь. Она хотела думать о другом, хотела молиться и ничего не могла сделать. Она большими открытыми глазами смотрела на лунный свет и тени, всякую секунду ждала увидеть его мертвое лицо и чувствовала, что тишина, стоявшая над домом и в доме, заковывала ее.
– Дуняша! – прошептала она. – Дуняша! – вскрикнула она диким голосом и, вырвавшись из тишины, побежала к девичьей, навстречу бегущим к ней няне и девушкам.
17 го августа Ростов и Ильин, сопутствуемые только что вернувшимся из плена Лаврушкой и вестовым гусаром, из своей стоянки Янково, в пятнадцати верстах от Богучарова, поехали кататься верхами – попробовать новую, купленную Ильиным лошадь и разузнать, нет ли в деревнях сена.
Богучарово находилось последние три дня между двумя неприятельскими армиями, так что так же легко мог зайти туда русский арьергард, как и французский авангард, и потому Ростов, как заботливый эскадронный командир, желал прежде французов воспользоваться тем провиантом, который оставался в Богучарове.
Ростов и Ильин были в самом веселом расположении духа. Дорогой в Богучарово, в княжеское именье с усадьбой, где они надеялись найти большую дворню и хорошеньких девушек, они то расспрашивали Лаврушку о Наполеоне и смеялись его рассказам, то перегонялись, пробуя лошадь Ильина.
Ростов и не знал и не думал, что эта деревня, в которую он ехал, была именье того самого Болконского, который был женихом его сестры.
Ростов с Ильиным в последний раз выпустили на перегонку лошадей в изволок перед Богучаровым, и Ростов, перегнавший Ильина, первый вскакал в улицу деревни Богучарова.
– Ты вперед взял, – говорил раскрасневшийся Ильин.
– Да, всё вперед, и на лугу вперед, и тут, – отвечал Ростов, поглаживая рукой своего взмылившегося донца.
– А я на французской, ваше сиятельство, – сзади говорил Лаврушка, называя французской свою упряжную клячу, – перегнал бы, да только срамить не хотел.
Они шагом подъехали к амбару, у которого стояла большая толпа мужиков.
Некоторые мужики сняли шапки, некоторые, не снимая шапок, смотрели на подъехавших. Два старые длинные мужика, с сморщенными лицами и редкими бородами, вышли из кабака и с улыбками, качаясь и распевая какую то нескладную песню, подошли к офицерам.
– Молодцы! – сказал, смеясь, Ростов. – Что, сено есть?
– И одинакие какие… – сказал Ильин.
– Развесе…oo…ооо…лая бесе… бесе… – распевали мужики с счастливыми улыбками.
Один мужик вышел из толпы и подошел к Ростову.
– Вы из каких будете? – спросил он.
– Французы, – отвечал, смеючись, Ильин. – Вот и Наполеон сам, – сказал он, указывая на Лаврушку.
– Стало быть, русские будете? – переспросил мужик.
– А много вашей силы тут? – спросил другой небольшой мужик, подходя к ним.
– Много, много, – отвечал Ростов. – Да вы что ж собрались тут? – прибавил он. – Праздник, что ль?
– Старички собрались, по мирскому делу, – отвечал мужик, отходя от него.
В это время по дороге от барского дома показались две женщины и человек в белой шляпе, шедшие к офицерам.
– В розовом моя, чур не отбивать! – сказал Ильин, заметив решительно подвигавшуюся к нему Дуняшу.
– Наша будет! – подмигнув, сказал Ильину Лаврушка.
– Что, моя красавица, нужно? – сказал Ильин, улыбаясь.
– Княжна приказали узнать, какого вы полка и ваши фамилии?
Пластиды – это мембранные органоиды, встречающиеся у фотосинтезирующих эукариотических организмов (высшие растения, низшие водоросли, некоторые одноклеточные организмы). У высших растений найден целый набор различных пластид (хлоропласт, лейкопласт, амилопласт, хромопласт), представляющих собой ряд взаимных превращений одного вида пластиды в другой. Основной структурой, которая осуществляет фотосинтетические процессы, является хлоропласт (рис. 226а).
Хлоропласт. Как уже указывалось, строение хлоропласта в принципе напоминает строение митохондрии. Обычно это структуры удлиненной формы с шириной 2-4 мкм и протяженностью 5-10 мкм. У зеленых водорослей встречаются гигантские хлоропласты (хроматофоры), достигающие длины 50 мкм. Количество хлоропластов в клетках растений различно. Так, у зеленых водорослей может быть по одному хлоропласту, у высших растений приходится в среднем 10-30 и в гигантских клетках палисадной ткани махорки обнаружено около 1000 хлоропластов на клетку.
Внешняя мембрана хлоропластов, как и внутренняя, имеют толщину около 7 мкм, они отделены друг от друга межмембранным пространством около 20-30 нм. Внутренняя мембрана хлоропластов отделяет строму пластиды, аналогичную матриксу митохондрий. В строме зрелого хлоропласта высших растений видны два типа внутренних мембран. Это – мембраны, образующие плоские, протяженные ламеллы стромы, и мембраны тилакоидов, плоских дисковидных вакуолей или мешков.
Ламеллы стромы (толщиной около 20 мкм) представляют собой плоские полые мешки или же имеют вид сети из разветвленных и связанных друг с другом каналов, располагающихся в одной плоскости. Обычно ламеллы стромы внутри хлоропласта лежат параллельно друг другу и не образуют связей между собой.
Кроме мембран стромы в хлоропластах имеются мембранные тилакоиды. Это плоские замкнутые мембранные мешки, имеющие форму диска. Величина межмембранного пространства у них также около 20-30 нм. Такие тилакоиды образуют стопки наподобие столбика монет, называемые гранами (рис. 227). Число тилакоидов на одну грану очень варьирует: от нескольких штук до 50 и более. Размер таких стопок может достигать 0,5 мкм, поэтому граны видны в некоторых объектах в световом микроскопе. Количество гран в хлоропластах высших растений может достигать 40-60. Тилакоиды в гране сближены друг с другом так, что внешние слои их мембран тесно соединяются; в месте соединения мембран тилакоидов образуется плотный слой толщиной около 2 нм. В состав граны кроме замкнутых камер тилакоидов обычно входят и участки ламелл, которые в местах контакта их мембран с мембранами тилакоидов тоже образуют плотные 2-нм слои. Ламеллы стромы, таким образом, как бы связывают между собой отдельные граны хлоропласта. Однако полости камер тилакоидов всезда замкнуты и не переходят в камеры межмембранного пространства ламелл стромы. Ламеллы стромы и мембраны тилакоидов образуются путем отделения от внутренней мембраны при начальных этапах развития пластид.
В матриксе (строме) хлоропластов содержатся молекулы ДНК, рибосомы; там же происходит первичное отложение запасного полисахарида, крахмала, в виде крахмальных зерен.
Функции хлоропластов. В хлоропластах происходят фотосинтетические процессы, приводящие к связыванию углекислоты, к выделению кислорода и синтезу сахаров.
Характерным для хлоропластов является наличие в них пигментов, хлорофиллов, которые и придают окраску зеленым растениям. С помощью хлорофилла зеленые растения поглощают энергию солнечного света и превращают ее в химическую.
Главным итоговым процессом здесь является связывание двуокиси углерода с использование воды для образования различных углеводов и выделение кислорода. Молекулы кислорода, который выделяется в процессе фотосинтеза у растений, образуется за счет гидролиза молекулы воды. Процесс фотосинтеза представляет собой сложную цепь, состоящую из двух фаз: световой и темновой. Первая, протекающая только на свету, связана с поглощением света хлорофиллами и с проведением фотохимической реакции (реакция Хилла). Во второй фазе, которая протекает в темноте, происходит фиксация и восстановление СО2 , приводящие к синтезу углеводов.
В результате световой фазы происходит синтез АТФ и восстановление НАДФ (никотинамидадениндинуклеотидфосфат), которые затем используются при восстановлении СО2, в синтезе углеводов уже в темновой фазе фотосинтеза.
В темновой стадии фотосинтеза за счет восстановленного НАДФ и энергии АТФ происходит связывание атмосферного СО2, что приводит к образованию углеводов. Этот процесс фиксации СО2 и образования углеводов состоит из многих этапов, в которых участвует большое число ферментов (цикл Кальвина).
В строме хлоропластов происходит восстановление нитритов до аммиака, за счет энергии электронов, активированных светом; в растениях этот аммиак служит источником азота при синтезе аминокислот и нуклеотидов.
Онтогенез и функциональные перестройки пластид. Увеличение числа хлоропластов и образование других форм пластид (лейкопластов и хромопластов) рассматривается как путь превращения структур-предшественников, пропластид . Весь же процесс развития различных пластид представляется идущего в одном направлении ряда смены форм:
Пропластида ® лейкопласт ® хлоропласт ® хромопласт
¯ амилопласт¾¾¾¾¾¾¾¾¾¾
Установлен необратимый характер онтогенетических переходов пластид. У высших растений возникновение и развитие хлоропластов происходят через изменения пропластид (рис. 231).
Пропластиды представляют собой мелкие (0,4-1 мкм) двумембранные пузырьки, отличающиеся от вакуолей цитоплазмы более плотным содержимым и наличием двух отграничивающих мембран, внешней и внутренней (наподобие промитохондриям у дрожжевых клеток). Внутренняя мембрана может давать небольшие складки или образовывать мелкие вакуоли. Пропластиды чаще всего встречаются в делящихся тканях растений (клетки меристемы корня, листьев, в точки роста стеблей и др.). Увеличение их числа происходит путем деления или почкования, отделения от тела пропластиды мелких двумембранных пузырьков.
Судьба таких пропластид зависит от условий развития растений. При нормальном освещении пропластиды превращаются в хлоропласты. Сначала они растут, при этом происходит образование продольно расположенных мембранных складок от внутренней мембраны. Одни из них простираются по всей длине пластиды и формируют ламеллы стромы; другие образуют ламеллы тилакоидов, которые выстраиваются в виде стопки и образуют граны зрелых хлоропластов.
В темноте у проростков происходит в начале увеличение объема пластид, этиопластов, но система внутренних мембран не строит ламеллярные структуры, а образует массу мелких пузырьков, которые скапливаютсяя в отдельные зоны и даже могут формировать сложные решетчатые структуры (проламеллярные тела). В мембранах этиопластов содержится протохлорофилл, предшественник хлорофилла желтого цвета. При освещении клеток мембранные пузырьки и трубочки быстро реорганизуются, из них развивается полная система ламелл и тилакоидов, характерная для нормального хлоропласта.
Лейкопласты в отличие от хлоропластов не имеют развитой ламеллярной системы (рис. 226 б). Встречаются они в клетках запасающих тканей. Из-за их неопределенной морфологии лейкопласты трудно отличить от пропластид, а иногда и от митохондрий. Они, как и пропластиды, бедны ламеллами, но тем не менее способны к образованию под влиянием света нормальных тилакоидных структур и к приобретению зеленой окраски. В темноте лейкопласты могут накапливать в проламеллярных телах различные запасные вещества, а в строме лейкопластов откладываются зерна вторичного крахмала. Если в хлоропластах происходит отложение так называемого транзиторного крахмала, который присутствует здесь лишь во время ассимиляции СО2, то в лейкопластах может происходить истинное запасание крахмала. В некоторых тканях (эндосперм злаков, корневища и клубни) накопление крахмала в лейкопластах приводит к образованию амилопластов, сплошь заполненных гранулами запасного крахмала, расположенных в строме пластиды (рис. 226в).
Другой формой пластид у высших растений является хромопласт, окрашивающийся обычно в желтый свет в результате накопления в нем каротиноидов (рис. 226г). Хромопласты образуются из хлоропластов и значительно реже их лейкопластов (например, в корне моркови). Процесс обесцвечивания и изменения хлоропластов легко наблюдать при развитии лепестков или при созревании плодов. При этом в пластидах могут накапливаться окрашенные в желтый цвет капельки (глобулы) или в них появляются тела в форме кристаллов. Эти процессы обусловлены постепенным уменьшением числа мембран в пластиде, с исчезновением хлорофилла и крахмала. Процесс образования окрашенных глобул объясняется тем, что при разрушении ламелл хлоропластов выделяются липидные капли, в которых хорошо растворяются различные пигменты (например, каротиноиды). Таким образом, хромопласты представляют собой дегенерирующие формы пластид, подвернутые липофанерозу – распаду липопротедных комплексов.
Фотосинтезирующие структуры низших эукариотических и прокариотических клеток . Строение пластид у низших фотосинтезирующих растений (зеленые, бурые и красные водоросли) в общих чертах сходно с хлоропластами клеток высших растений. Их мембранные системы также содержат фоточувствительные пигменты. Хлоропласты зеленых и бурых водорослей (иногда их называют хроматофорами) имеют также внешнюю и внутреннюю мембраны; последняя образует плоские мешки, располагающиеся параллельными слоями, граны у этих форм не встречаются (рис. 232). У зеленых водорослей в состав хроматофора входят пиреноиды , представляющие собой окруженные мелкими вакуолями зону, вокруг которой происходит отложение крахмала (рис. 233).
Форма хлоропластов у зеленых водорослей очень разнообразна – это или длинные спиральные ленты (Spirogira), сети (Oedogonium), или мелкие округлые, похожие на хлоропласты высших растений (рис. 234).
Среди прокариотических организмов многие группы обладают фотосинтетическими аппаратами и имеют в связи с этим особое строение. Для фотосинтезирующих микроорганизмов (синезеленые водоросли и многие бактерии) характерно, что их фоточувствительные пигменты связаны с плазматической мембраной или с её выростами, направленными вглубь клетки.
В мембранах синезеленых водорослей кроме хлорофилла находятся пигменты фикобилины. Фотосинтезирующие мембраны синезеленых водорослей образуют плоские мешки (ламеллы), которые располагаются параллельно друг над другом, иногда образуя стопки или спирали. Все эти мембранные структуры образуются за счет инвагинаций плазматической мембраны.
У фотосинтезирующих бактерий (Chromatium) мембраны образуют мелкие пузырьки, число которых так велико, что они заполняют практически большую часть цитоплазмы.
Геном пластид. Подобно митохондриям, хлоропласты имеют собственную генетическую систему, обеспечивающую синтез ряда белков внутри самих пластид. В матриксе хлоропластов обнаруживаются ДНК, разные РНК и рибосомы. Оказалось, что ДНК хлоропластов резко отличается от ДНК ядра. Она представлена циклическими молекулами длиной до 40-60 мкм, имеющими молекулярный вес 0,8-1,3х108 дальтон. В одном хлоропласте может быть множество копий ДНК. Так, в индивидуальном хлоропласте кукурузы присутствует 20-40 копий молекул ДНК. Длительность цикла и скорость репликации ядерной и хлоропластной ДНК, как было показано на клетках зеленых водорослей, не совпадают. ДНК хлоропластов не состоит в комплексе с гистонами. Все эти характеристики ДНК хлоропластов близки к характеристикам ДНК прокариотических клеток. Более того, сходство ДНК хлоропластов и бактерий подкрепляется еще и тем, что основные регуляторные последовательности транскрипции (промоторы, терминаторы) у них одинаковы. На ДНК хлоропластов синтезируются все виды РНК (информационная, трансферная, рибосомная). ДНК хлоропластов кодирует рРНК, входящую в состав рибосом этих пластид, которые относятся к прокариотическому 70S типу (содержат 16S и 23S рРНК). Рибосомы хлоропластов чувствительны к антибиотику хлорамфениколу, подавляющему синтез белка у прокариотических клеток.
Полностью расшифрована вся последовательность нуклеотидов в составе циклической молекулы ДНК хлоропластов высших растений. Эта ДНК может кодировать до 120 генов, среди них: гены 4 рибосомных РНК, 20 рибосомных белков хлоропластов, гены некоторых субъединиц РНК-полимеразы хлоропластов, несколько белков I и II фотосистем, 9 из 12 субъединиц АТФ-синтетазы, части белков комплексов цепи переноса электронов, одной из субъединиц рибулозодифосфат-карбоксилазы (ключевой фермент связывания СО2), 30 молекул тРНК и еще 40 пока неизвестных белков. Интересно, что сходный набор генов в ДНК хлоропластов обнаружен у таких далеко отстоящих представителей высших растений как табак и печеночный мох.
Основная же масса белков хлоропластов контролируется ядерным геномом. Ряд важнейших белков, ферментов, а соответственно и метаболические процессы хлоропластов находятся под генетическим контролем ядра. Под контролем ядерных генов находится большая часть рибосомных белков. Все эти данные говорят о хлоропластах, как о структурах с ограниченной автономией.
4.6. Цитоплазма: Опорно-двигательная система (цитоскелет)
В основе всех многочисленных двигательных реакций клетки лежат общие молекулярные механизмы. Кроме того, наличие двигательных аппаратов сочетается и структурно связано с существованием опорных, каркасных или скелетных внутриклеточных образований. Поэтому говорят об опорно-двигательной системе клеток.
К цитоскелетным компоненты относятся нитевидные, неветвящиеся белковые комплексы или филаменты (тонкие нити).
Существуют три группы филаментов, различающихся как по химическому составу и ультраструктуре, так и по функциональным свойствам. Самые тонкие нити – это микрофиламенты; их диаметр составляет около 8 нм и состоят они в основном из белка актина. Другую группу нитчатых структур составляют микротрубочки, которые имеют диаметр 25 нм и состоят в основном из белка тубулина, и, наконец, промежуточные филаменты с диметром около 10 нм (промежуточный по сравнению с 6 нм и 25 нм), образующиеся из разных, но родственных белков (рис. 238, 239).
Все эти фибриллярные структуры участвуют в процессах физического перемещения клеточных компонентов или даже целых клеток, в ряде случаев выполняют сугубо каркасную скелетную роль. Элементы цитоскелета встречаются во всех без исключения эукариотических клетках; аналоги этих фибриллярных структур встречаются и у прокариот.
Общими свойствами элементов цитоскелета является то, что это белковые, неветвящиеся фибриллярные полимеры, нестабильные, способные к полимеризации и деполимеризации, которые приводят к клеточной подвижности, например, к изменению формы клетки. Компоненты цитоскелета при участии специальных дополнительных белков могут стабилизироваться или образовывать сложные фибриллярные ансамбли, и играть только каркасную роль. При взаимодействии с другими специальными белками-транслокаторами (или моторными белками) они участвуют в разнообразных клеточных движениях.
По своим свойствам и функциям элементы цитоскелета разделяют на две группы: только каркасные фибриллы – промежуточные филаменты, и опорно-двигательные – актиновые микрофиламенты, взаимодействующие с моторными белками – миозинами, и тубулиновые микротрубочки, взаимодействующие с моторными белками динеинами и кинезинами.
Вторая группа фибрилл цитоскелета (микрофиламенты и микротрубочки) обеспечивают два принципиально различных способа движения. Первый из них основан на способности основного белка микрофиламентов – актина и основного белка микротрубочек – тубулина к полимеризации и деполимеризации. При связи этих белков с плазматической мембраной наблюдаются ее морфологические изменения в виде образования выростов (псевдоподий и ламеллоподий) на краю клетки.
При другом способе передвижения фибриллы актина (микрофиламенты) или тубулина (микротрубочки) являются направляющими структурами, по которым перемещаются специальные подвижные белки - моторы. Последние могут связываться с мембранными или фибриллярными компонентами клетки и тем самым участвовать в их перемещении.