Прием-прием, есть кто? Азбука Бодибилдинга на связи! И в этот пятничный денек мы разберем необычную тему под названием электрическая активность мышц.
По прочтении Вы узнаете что такое ЭМГ как явление, для чего и в каких целях используется данный процесс, почему большинство исследований по “лучшести” упражнений оперируют именно данными электрической активности.
![]()
Итак, рассаживайтесь поудобней, будет интересно.
Электрическая активность мышц: вопросы и ответы
Эта уже вторая по счету статья в цикле “Muscle inside”, в первой мы говорили про , а в целом цикл посвящен явлениям и мероприятиям, которые протекают (могут протекать) внутри мускулов. Данные заметки позволят Вам лучше понимать накачательные процессы и быстрее прогрессировать в улучшении телосложения. Почему мы, собственно, решили рассказать именно про электрическую активность мышц? Все очень просто. В наших технических (и не только) статьях мы постоянно приводим списки из лучших упражнений, которые формируются именно на основании данных исследований по ЭМГ.
Вот уже на протяжении практически пяти лет, мы сообщаем Вам эту информацию, но ни разу за это время не раскрыли саму суть явления. Что же, сегодня мы восполним этот пробел.
Примечание:
Все дальнейшее повествование по теме электрическая активность мышц, будет разбито на подглавы.
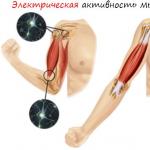
Что такое электромиография? Замер активности мышц
ЭМГ представляет собой метод электродиагностической медицины для оценки и регистрации электрической активности, создаваемой скелетными мышцами. Процедура ЭМГ выполняется с использованием прибора, называемого электромиографом, для создания записи, называемой электромиограммой. Электромиограф обнаруживает электрический потенциал, генерируемый мышечными клетками, когда они электрически или неврологически активированы. Для понимания сути явления ЭМГ необходимо иметь представление о структуре мышц и протекающих внутри процессах.
Мышца представляет собой организованную “коллекцию” мышечных волокон (м.в.) , которые, в свою очередь, состоят из групп компонентов, известных как миофибриллы. В костно-скелетной системе нервные волокна инициируют электрические импульсы в м.в., известные как потенциалы действия мышц. Они создают химические взаимодействия, которые активируют сокращение миофибрилл. Чем больше активированных волокон в мышечной части, тем сильнее сокращение, которое может произвести мускул. Мышцы могут только создавать усилие при своем сокращении/укорочении. Тяговое и толкающее усилие в костно-мышечной системе генерируется сопряжением мышц, которые действуют в антагонистической модели: одна мышца сокращается, а другая расслабляется. Например, при подъеме гантели на бицепс, двуглавая мышца плеча при подъеме снаряда сокращается/укорачивается, а трицепс (антагонист) находится в расслабленном состоянии.

ЭМГ в различных видах спорта
Метод оценки основной мышечной активности, возникающей при физическом движении, получил широкое распространение во многих видах спорта, особенно фитнесе и бодибилдинге. Измеряя количество и величину импульсов, возникающих во время мышечной активации, можно оценить, насколько стимулируется мышечная единица, чтобы придать особую силу. Электромиограмма представляет собой визуальную иллюстрацию сигналов, генерируемых во время мышечной активности. И далее по тексту мы рассмотрим некоторые “портреты” ЭМГ.
Процедура ЭМГ. Из чего она состоит и где проводится?
В большинстве своем замерить электрическую активность мышц возможно только в специальных научно-исследовательских спортивных лабораториях, т.е. профильных учреждениях. Современные фитнес-клубы не предоставляют такой возможности ввиду отсутствия квалифицированных специалистов и низкой востребованности со стороны аудитории клуба.
Сама процедура состоит из:
- размещения на теле человека в определенной области (на или рядом с исследуемой мышечной группой) специальных электродов, подсоединенных к блоку, измеряющему электрические импульсы;
- запись и передача сигналов в компьютер через блок беспроводной передачи данных ЭМГ от расположенных поверхностных электродов для последующего отображения и анализа.
В картинном варианте процедура ЭМГ выглядит следующим образом.

Мышечная ткань в состоянии покоя электрически неактивна. Когда мышца добровольно сжимается, начинают появляться потенциалы действия. По мере увеличения силы сокращения мышц все больше и больше мышечных волокон вырабатывают потенциалы действия. Когда мышца полностью сжимается, должна появиться беспорядочная группа потенциалов действия с различными скоростями и амплитудами (полный набор и интерференционная картина) .
Таким образом, процесс получения картинки сводится к тому, что испытуемый выполняет конкретное упражнение по конкретной схеме (сеты/повторения/отдых) , а приборы фиксируют генерируемые мышцами электрические импульсы. В конечном итоге результаты отображаются на экране ПК в виде определенного графика импульсов.
Чистота результатов ЭМГ и понятие MVC
Как Вы, наверное, помните из наших технических заметок, иногда мы приводили разные значения по электрической активности мышц даже для одного и того же упражнения. Это связано с тонкостями проведения самой процедуры. В целом на конечные результаты оказывает влияние ряд факторов:
- выбор конкретной мышцы;
- размер самой мышцы (у мужчин и женщин разные объемы) ;
- правильное размещение электрода (в конкретном месте поверхностной мышцы – брюшко мышцы, продольная средняя линия) ;
- процент жира в организме человека (чем больше жира, тем слабее сигнал ЭМГ) ;
- толщина – насколько сильно ЦНС генерирует сигнал, насколько быстро он поступает в мускул;
- стаж тренировок – насколько у человека хорошо развита .
Таким образом, ввиду указанных начальных условий разные исследования могут давать разные результаты.
Примечание:
Более точные результаты активности мышц в конкретном движении дает внутримышечный метод оценки. Это когда игольчатый электрод вводят через кожу в мышечную ткань. Игла затем перемещается в несколько точек в расслабленной мышце, чтобы оценить как активность вставки, так и активность покоя в мышцах. Оценивая активность покоя и вставки, электромиограф оценивает активность мышц во время добровольного сокращения. По форме, размеру и частоте результирующих электрических сигналов судят о степени активности конкретной мышцы.
В процедуре электромиографии одной из основных ее функций является то, как хорошо можно активировать мышцу. Наиболее распространенный способ это выполнение максимального добровольного сокращения (MVC) тестируемой мышцы. Именно MVC, в большинстве исследований, принимается как наиболее достоверное средство анализа пиковой силы и силы, создаваемой мышцами.
Однако наиболее полную картину по активности мышц способно дать предоставление обоих наборов данных (MVC и ARV – средние) значений ЭМГ.
Собственно, с теоретической частью заметки разобрались, теперь окунемся в практику.
Электрическая активность мышц: лучшие упражнения для каждой мышечной группы, результаты исследований
Сейчас мы начнем собирать шишки:) от нашей многоуважаемой аудитории, и все потому, что займемся неблагодарным занятием – доказыванием того, что конкретное упражнение является лучшим для конкретной мышечной группы.
А почему оно неблагодарное, Вы поймете по ходу повествования.
Итак, принимая показания ЭМГ во время различных упражнений, мы можем нарисовать иллюстративную картину уровня активности и возбуждения внутри мышцы. Это может указывать, насколько эффективным является конкретное упражнение при стимуляции конкретного мускула.
I. Результаты исследований (профессор Tudor Bompa, Mauro Di Pasquale, Италия 2014)
Данные представлены по шаблону, мышечная группа-упражнение-процент активации м.в.:
Примечание:
Процентное значение указывает пропорцию активированных волокон, значение 100% означает полную активацию.
№1. Широчайшие мышцы спины:
- – 91 ;
- – 89 ;
- – 86 ;
- – 83 .
№2. Грудные мышцы (большая пекторальная) :
- – 93 ;
- – 87 ;
- – 85 ;
- – 84 .
№3. Передняя дельта:
- жим гантелей стоя – 79 ;
- – 73 .
№4. Средняя/боковая дельта:
- подъемы прямых рук через стороны с гантелями – 63 ;
- подъемы прямых рук через стороны на верхнем блоке кроссовера – 47 .
№5. Задняя дельта:
- разведение рук в наклоне стоя с гантелями - 85 ;
- разведение рук в наклоне стоя с нижнего блока кроссовера – 77 .
№6. Бицепс (длинная головка) :
- сгибание рук на скамье Скотта с гантелями – 90 ;
- сгибание рук с гантелями сидя на скамье под углом вверх - 88 ;
- (узкий хват) – 86 ;
- – 84 ;
- – 80 .
№7. Квадрицепс (прямая мышца бедра) :
- – 88 ;
- – 86 ;
- – 78 ;
- – 76 .
№8. Задняя поверхность (бицепс) бедра:
- – 82 ;
- – 56 .
№9. Задняя поверхность (полусухожильная мышца) бедра:
- – 88 ;
- становая тяга на прямых ногах – 63 .
С уважением и признательностью, Протасов Дмитрий
.
Мышечную систему образно определяют как биологический ключ человека к внешнему миру.
Электромиография
- метод исследования функционального состояния органов движения путем регистрации биопотенциалов мышц
. Электромиография - это регистрация электрических процессов в мышцах, фактически запись потенциалов действия мышечных волокон, которые заставляют ее сокращаться. Мышца представляет собой массу ткани, состоящую из множества отдельных мышечных волокон, соединенных вместе и работающих согласованно. Каждое мышечное волокно - это тонкая нить, толщиной всего лишь около 0,1 мм до 300 мм длиной. При стимуляции электрическим потенциалом действия, приходящим к волокну от мотонейрона, это волокно сокращается иногда примерно до половины первоначальной длины. Мышцы, участвующие в тонких двигательных коррекциях (фиксация объекта глазами), могут иметь в каждой единице всего по 10 волокон. В мышцах, осуществляющих более грубую регулировку при поддержании позы, в одной двигательной единице может быть до 3000 мышечных волокон. Поверхностная электромиограмма (ЭМГ) суммарно отражает разряды двигательных единиц, вызывающих сокращение. Регистрация ЭМГ позволяет выявить намерение начать движение за несколько секунд до его реального начала. Помимо этого миограмма выступает как индикатор мышечного напряжения. В состоянии относительного покоя связь между действительной силой, развиваемой мышцей, и ЭМГ линейна.
Поверхностная электромиограмма (ЭМГ) суммарно отражает разряды двигательных единиц, вызывающих сокращение. Регистрация ЭМГ позволяет выявить намерение начать движение за несколько секунд до его реального начала. Помимо этого миограмма выступает как индикатор мышечного напряжения. В состоянии относительного покоя связь между действительной силой, развиваемой мышцей, и ЭМГ линейна.
Прибор, с помощью которого регистрируются биопотенциалы мышц, называется электромиографом, а регистрируемая с его помощью запись электромиограммой (ЭМГ). ЭМГ, в отличие от биоэлектрической активности мозга (ЭЭГ), состоит из высокочастотных разрядов мышечных волокон, для неискаженной записи которых, по некоторым представлениям, требуется полоса пропускания до 10 000 Гц.
Показатели активности дыхательной системы
 Дыхательная система состоит из дыхательных путей и легких.
Дыхательная система состоит из дыхательных путей и легких.
Основной двигательный аппарат этой системы составляют межреберные мышцы, диафрагма и мышцы живота. Воздух, поступающий в легкие во время вдоха, снабжает протекающую по легочным капиллярам кровь кислородом. Одновременно из крови выходят двуокись углерода и другие вредные продукты метаболизма, которые выводятся наружу при выдохе. Между интенсивностью мышечной работы, совершаемой человеком, и потреблением кислорода существует простая линейная зависимость.
В психофизиологических экспериментах в настоящее время дыхание регистрируется относительно редко, главными образом для того, чтобы контролировать артефакты. Для измерения интенсивности (амплитуды и частоты) дыхания используют специальный прибор - пневмограф. Он состоит из надувной камеры-пояса, плотно оборачиваемой вокруг грудной клетки испытуемого, и отводящей трубки, соединенной с манометром и регистрирующим устройством. Возможны и другие способы регистрации дыхательных движений, но в любом случае обязательно должны присутствовать датчики натяжения, фиксирующие изменение объема грудной клетки.
Для измерения интенсивности (амплитуды и частоты) дыхания используют специальный прибор - пневмограф. Он состоит из надувной камеры-пояса, плотно оборачиваемой вокруг грудной клетки испытуемого, и отводящей трубки, соединенной с манометром и регистрирующим устройством. Возможны и другие способы регистрации дыхательных движений, но в любом случае обязательно должны присутствовать датчики натяжения, фиксирующие изменение объема грудной клетки.
Этот метод обеспечивает хорошую запись изменений частоты и амплитуды дыхания. По такой записи легко анализировать число вдохов в минуту, а также амплитуду дыхательных движений в разных условиях. Можно сказать, что дыхание - это один из недостаточно оцененных факторов в психофизиологических исследованиях.
Реакции глаз
Для психофизиолога наибольший интерес представляют три категории глазных реакций: сужение и расширение зрачка, мигание и глазные движения.
Пупиллометрия
- метод изучения зрачковых реакций.
Зрачок - отверстие в радужной оболочке, через которое свет попадает на сетчатку. Диаметр зрачка человека может меняться в пределах от 1,5 до 9 мм. Величина зрачка существенно колеблется в зависимости от количества света, падающего на глаз: на свету зрачок сужается, в темноте - расширяется. Наряду с этим, размер зрачка существенно изменяется, если испытуемый реагирует на воздействие эмоционально. В связи с этим пупиллометрия используется для изучения субъективного отношения людей к тем или иным внешним раздражителям.
Диаметр зрачка можно измерять путем простого фотографирования глаза в ходе обследования или же с помощью специальных устройств, преобразующих величину зрачка в постоянно варьирующий уровень потенциала, регистрируемый на полиграфе.
Мигание
(моргание) - периодическое смыкание век
. Длительность одного мигания приблизительно 0,35 с. Средняя частота мигания составляет 7,5 в минуту и может варьировать в пределах от 1 до 46 в минуту. Мигание выполняет разные функции в обеспечении жизнедеятельности глаз. Однако для психофизиолога существенно, что частота мигания изменяется в зависимости от психического состояния человека.
Движение глаз
широко исследуются в психологии и психофизиологии. Это разнообразные по функции, механизму и биомеханике вращения глаз в орбитах. Существуют разные типы глазных движений, выполняющие различные функции. Однако наиболее важная среди них функция движений глаз состоит в том, чтобы поддерживать интересующее человека изображение в центре сетчатки, где самая высокая острота зрения. Минимальная скорость прослеживающих движений около 5 угл. мин/с, максимальная достигает 40 град/с.
Электроокулография
- метод регистрации движения глаз
, основанный на графической регистрации изменения электрического потенциала сетчатки и глазных мышц. У человека передний полюс глаза электрически положителен, а задний отрицателен, поэтому существует разность потенциалов между дном глаза и роговицей, которую можно измерить. При повороте глаза положение полюсов меняется, возникающая при этом разность потенциалов характеризует направление, амплитуду и скорость движения глаза. Это изменение, зарегистрированное графически, носит название электроокулограммы. Однако микродвижения глаз с помощью этого метода не регистрируются, для их регистрации разработаны другие приемы. (см. рис.)
Детектор лжи
Детектор лжи
- условное название прибора полиграфа, одновременно регистрирующего комплекс физиологических показателей (КГР, ЭЭГ, плетизмограмму и др.) с целью выявить динамику эмоционального напряжения. С человеком, проходящем обследование на полиграфе, проводят собеседование, в ходе которого наряду с нейтральными задают вопросы, составляющие предмет специальной заинтересованности. По характеру физиологических реакций, сопровождающих ответы на разные вопросы, можно судить об эмоциональной реактивности человека и в какой-то мере о степени его искренности в данной ситуации. Поскольку в большинстве случаев специально необученный человек не контролирует свои вегетативные реакции, детектор лжи дает по некоторым оценкам до 71% случаев обнаружения обмана.
Следует иметь в виду, однако, что сама процедура собеседования (допроса) может быть настолько неприятна для человека, что возникающие по ходу физиологические сдвиги будут отражать эмоциональную реакцию человека на процедуру. Отличить спровоцированные процедурой тестирования эмоции от эмоций, вызванных целевыми вопросам, невозможно. В то же время человек, обладающий высокой эмоциональной стабильностью, сможет относительно спокойно чувствовать себя в этой ситуации, и его вегетативные реакции не дадут твердых основания для вынесения однозначного суждения. По этой причине к результатам, полученных с помощью детектора лжи, нужно относиться с должной мерой критичности (см. Видео).
Выбор методик и показателей
В идеале выбор физиологических методик и показателей должен логически вытекать из принятого исследователем методологического подхода и целей, поставленных перед экспериментом. Однако на практике нередко исходят из других соображений, например, доступности приборов и легкости обработки экспериментальных данных.
Более весомыми представляются аргументы в пользу выбора методик, если извлекаемые с их помощью показатели получают логически непротиворечивое содержательное толкование в контексте изучаемой психологической или психофизиологической модели.
Психофизиологические модели.
В науке под моделью понимается упрощенное знание, несущее определенную, ограниченную информацию об объекте/явлении, отражающее те или иные его свойства. С помощью моделей можно имитировать функционирование и прогнозировать свойства изучаемых объектов, процессов или явлений. В психологии моделирование имеет два аспекта: моделирование психики
и моделирование ситуаций
. Под первым подразумевается знаковая или техническая имитация механизмов, процессов и результатов психической деятельности, под вторым организация того или иного вида человеческой деятельности путем искусственного конструирования среды, в которой осуществляется эта деятельность.
Оба аспекта моделирования находят место в психофизиологических исследованиях. В первом случае моделируемые особенности деятельности человека, психических процессов и состояний прогнозируются на основе объективных физиологических показателей, нередко зарегистрированных вне прямой связи с изучаемым феноменом. Например, показано, что некоторые индивидуальные особенности восприятия и памяти можно прогнозировать по характеристикам биотоков мозга. Во втором случае психофизиологическое моделирование включает имитацию в лабораторных условиях определенной психической деятельности, с целью выявления ее физиологических коррелятов и /или механизмов. Обязательным при этом является создание некоторых искусственных ситуаций, в которых так или иначе включаются исследуемые психические процессы и функции. Примером такого подхода служат многочисленные эксперименты по выявлению физиологических Коррелятов восприятия, памяти и т.д.
При интерпретации результатов в подобных экспериментах исследователь должен четко представлять себе, что модель никогда не бывает полностью идентична изучаемому явлению или процессу. Как правило, в ней учитываются лишь какие-то отдельные стороны реальности. Следовательно, каким бы исчерпывающим ни казался, например, какой-либо психофизиологический эксперимент по выявлению нейрофизиологических коррелятов процессов памяти, он будет давать лишь частичное знание о природе ее физиологических механизмов, ограниченное рамками данной модели и используемых методических приемов и показателей. Именно по этой причине психофизиология изобилует разнообразием несвязанных между собой, а иногда и просто противоречивых экспериментальных данных. Полученные в контексте разных моделей такие данные представляют фрагментарное знание, которое в перспективе, вероятно, должно объединиться в целостную систему, описывающую механизмы психофизиологического функционирования.
Интерпретация показателей.
Особого внимания заслуживает вопрос о том, какое значение экспериментатор придает каждому из используемых им показателей. В принципе физиологические показатели могут выполнять две основные роли: целевую (смысловую) и служебную (вспомогательную). Например, при изучении биотоков мозга в процессе умственной деятельности целесообразно параллельно регистрировать движения глаз, мышечное напряжение и некоторые другие показатели. Причем в контексте такой работы только показатели биотоков мозга несут смысловую нагрузку, связанную с данной задачей. Остальные показатели служат для контроля артефактов и качества регистрации биотоков (регистрация глазных движений), контроля эмоциональных состояний испытуемого (регистрация КГР), поскольку, хорошо известно, что глазные движения и эмоциональное напряжение могут привносить помехи и искажать картину биотоков, особенно когда испытуемый решает какую-либо задачу. В то же время в другом исследовании регистрация и глазных движений, и КГР может играть смысловую, а не служебную роль. Например, когда предмет исследования - стратегия визуального поиска или изучение физиологических механизмов эмоциональной сферы человека.
Таким образом, один и тот же физиологический показатель может быть использован для решения разных задач. Другими словами, специфика использования показателя определяется не только его собственными функциональными возможностями, но также и тем психологическим контекстом, в который он включается. Хорошее знание природы и всех возможностей используемых физиологических показателей - важный фактор в организации психофизиологического эксперимента.
Значение экспериментов, выполненных на животных.
Как уже отмечалось выше, многие задачи в психофизиологии решались и продолжают решаться в экспериментах на животных. (В первую очередь речь идет об изучении активности нейронов.) В связи с этим особое значение приобретает проблема, сформулированная еще Л.С. Выготским. Это проблема специфического для человека соотношения структурных и функциональных единиц в деятельности мозга и определения новых по сравнению с животными принципов функционирования систем, внутри- и межсистемных взаимодействий.
Следует прямо указать, что проблема "специфического для человека соотношения структурных и функциональных единиц в деятельности мозга и определения новых по сравнению с животными" принципов функционирования систем, к сожалению, пока не получила продуктивного развития. Как пишет О.С. Андрианов (1993): "Стремительное "погружение" биологии и медицины... в глубины живой материи отодвинуло на задний план изучение важнейшей проблемы - эволюционной специфики мозга человека. Попытки найти на молекулярном уровне некий материальный субстрат, характерный только для мозга человека и определяющий особенности наиболее сложных психических функций, пока не увенчались успехом".
Таким образом, встает вопрос о правомерности переноса данных полученных на животных для объяснения мозговых функций у человека. Широко принята точка зрения, в соответствии с которой существуют универсальные механизмы клеточного функционирования и общие принципы кодирования информации, что позволяет осуществлять интерполяцию результатов (см., например: Основы психофизиологии под ред. Ю.И. Александрова, 1998).
Один из основателей отечественной психофизиологии Е.Н. Соколов, решая проблему переноса результатов исследований, выполненных на животных, на человека, сформулировал принцип психофизиологического исследования следующим образом: человек - нейрон - модель. Это значит, что психофизиологическое исследование начинается с изучения поведенческих (психофизиологических) реакций человека, Затем оно переходит к изучению механизмов поведения с помощью микроэлектродной регистрации нейронной активности в опытах на животных, а у человека - с использованием электроэнцефалограммы и вызванных потенциалов. Интеграция всех данных осуществляется путем построения модели из нейроподобных элементов. При этом вся модель как целое должна воспроизводить исследуемую функцию, а отдельные нейроподобные элементы должны обладать характеристиками и свойствами реальных нейронов. Перспективы исследований такого рода заключаются в построении моделей "специфически человеческого типа" таких, например, как нейроинтеллект.
Заключение.
Приведенные выше материалы свидетельствуют о большом разнообразии и разноуровневости психофизиологических методов. В сферу компетентности психофизиолога входит многое, начиная от динамики нейрональной активности в глубоких структурах мозга до локального кровотока в пальце руки. Закономерно возникает вопрос, каким образом объединить столь различные по способам получения и содержанию показатели в логически непротиворечивую систему. Решение его, однако, упирается в отсутствие единой общепринятой психофизиологической теории.
Психофизиология, которая родилась как экспериментальная ветвь психологии, в значительной степени остается таковой и по сей день, компенсируя несовершенство теоретического фундамента многообразием и изощренностью методического арсенала. Богатство этого арсенала велико, его ресурсы и перспективы представляются неисчерпаемыми. Стремительный рост новых технологий неизбежно расширит возможности проникновению в тайны человеческой телесности. Он приведет к созданию новых обрабатывающих устройств, способных формализовать сложную систему зависимости переменных величин, используемых в объективных физиологических показателях, закономерно связанных с психической деятельностью человека. Независимо от того, будут ли новые решения результатом дальнейшего развития электронно-вычислительной техники, эвристических моделей или других, еще неизвестных нам способов познания, развитие науки в наше время предвосхищает коренное преобразование психофизиологического мышления и методов работы
Словарь терминов
- альфа-ритм
- пейсмекер
- ретикулярная формация
- афферентация
- кортико-лимбическое взаимодействие
- кожно-гальваническая реакция (КГР)
Вопросы для самопроверки
- Как связаны ритмические составляющие электроэнцефалограммы с состоянием человека?
- Чем обусловлена кожно-гальваническая реакция?
- Как различаются пневмография и спирография?
- Что дает оценка состояния периферических сосудов?
- Как интерпретируют показатели детектора лжи?
Список литературы
- Анохин П.К. Очерки по физиологии функциональных систем. М.: Медицина, 1975.
- Буреш Я., Бурешова О., Хьюстон Д.П. Методики и основные эксперименты по изучению мозга и поведения. М.: Высшая школа, 1991.
- Беленков Н.Ю. Принцип целостности в деятельности мозга. М.: Медицина, 1980.
- Бернштейн Н.А. Очерки по физиологии движений и по физиологии активности. М.: Медицина, 1966.
- Бехтерева Н.П., Бундзен П.В., Гоголицын Ю.Л. Мозговые коды психической деятельности. Л.: Наука, 1977.
- Гнездицкий В.В. Вызванные потенциалы мозга в клинической практике. Таганрог: ТГТУ, 1997.
- Данилова Н.Н. Психофизиология. М.: Аспект Пресс, 1998.
- Дубровский Д.И. психика и мозг: результаты и перспективы исследований // Психологический журнал. 1990. Т.11. № 6. С. 3-15.
- Естественнонаучные основы психологии / Под. ред. А.А. Смирнова, А.Р. Лурия, В.Д. Небылицына. М.: Педагогика, 1978.
- Иваницкий А.М., Стрелец В.Б., Корсаков И.А. Информационные процессы мозга и психическая деятельность. М.: Наука, 1984.
- Ломов Б.Ф. методологические и теоретические проблемы психологии. М.: Наука, 1984.
- Нейрокомпьютер как основа мыслящих ЭВМ. М.: Наука, 1993.
- Мерлин В.С. Очерк интегрального исследования индивидуальности. М.: Педагогика, 1986.
- Методика и техника психофизиологического эксперимента. М.: Наука, 1987.
- Основы психофизиологии / Под ред. Ю.И. Александрова. М., 1998.
- Тихомиров О.К. Психология мышления. М.: МГУ, 1984.
- Чуприкова Н.И. психика и сознание как функция мозга. М.: Наука, 1985.
- Хэссет Дж. Введение в психофизиологию. М.: Мир, 1981.
- Ярвилехто Т. Мозг и психика. М.: Прогресс, 1992.
Вам никогда не приходила в голову такая мысль почему абсолютно все новички приходя в спортзал, в первый год своих тренировок не могут прибавить в мышечной массе? Когда-то и меня посетила эта самая мысль и я попытался в ней более подробно разобраться. И понял что на самом деле всё довольно-таки просто и что всё дело оказывается в нашей с вами мышечной активности.
Что такое мышечная активность?
Мышечная активность -это по большому счёту не что иное как ваше взаимодействие между используемой силой ваших мышц на данный момент с их максимальной возможностью.
Объясню вам немного проще. В обычной жизни мы как правило не используем силу наших мышц на полную мощность. И поэтому всё сводиться к тому, что человек вне спортзала в повседневной жизни работает и поднимает максимум, ну скажем 5 – 10 кг да и то только тогда, когда он идёт за продуктами в магазин.
Именно к такой нагрузке и привыкли мышцы обычного человека, который не ходит в спортзал и не поднимает железо…
Поэтому наши мышцы давно адаптировались к такой минимальной нагрузке и включаются не на полную мощность, а от силы на 5% — 10% процентов.
А вот их силовая возможность значительно выше, но при всём при этом они не будут включаться в работу просто так потому что вы этого захотели. Для этого их нужно тренировать.
Мышечная активность не тренированного человека составляет примерно 30% — 40% процентов это более чем в 2-3 раза ниже силовых возможностей и потенциала самих мышц.
Если не тренированный человек приложит максимум своих усилий к чему-либо, тогда его нынешняя максимальная мышечная активность будет составлять приблизительно 30% – 40% процентов, по сути это даже меньше половины от возможности самих мышц.
И вот когда такой человек решается пойти в спортзал для того чтобы начать активно заниматься. И после 3-4 месяцев он видит, что особо мышц он не прибавил за всё это время, но при этом он значительно прибавил в силе, то тут возникает резонный вопрос почему?
Ответ заключается в том, что он просто не прибавил в мышечной массе потому, что он не выработал достаточный потенциал своих мышц на данный момент времени.
Все мы давно с вами знаем что мышечный рост на прямую зависит от увеличения силы, поэтому чем сильнее будут ваши мышцы, тем больше они будут в объёме.
И это давно проверенный факт!
Мышечная масса, а точнее гипертрофия мышц происходит лишь тогда, когда мышцы испытывают сильнейшую нагрузку, а точнее даже сказать огромный мышечный перегруз.
А что мы видим обычно на практике? Так это то, что все новички в основном сидят на тренажёрах, да и как правило начинают качать самые ходовые мышцы, такие как бицепс, трицепс и пресс, иногда делая жим лёжа.
- Они приседают – НЕТ
- Они делают становую тягу – НЕТ
- Они делают жим стоя – тоже НЕТ
Большинство новичков в спортзале вообще не делают тяжёлых базовых упражнений. Они не работают тяжело и не выполняют действительно тяжёлые базовые упражнения, а выполняют как правило, всегда изолированные упражнения работая при этом в основном на тренажёрах.
И самое главное они даже не пытаются как-то увеличить свои силовые показатели, а лишь постоянно изнуряют свои мышцы малыми весами и большим количеством повторений.
Иными словами их мышечный потенциал не исчерпал своих силовых возможностей для их дальнейшего роста на данный момент. А это значит, что на данный момент вся их мышечная активность будет ниже 100% процентов.
Поэтому многие атлеты, а это почти 90% процентов даже через год и даже два года не видят никакого роста в мышечной массе…
Для того чтобы произошёл какой-то мышечный рост, на начальном этапе необходимо сначала увеличить силу своих мышц, а этот факт понимают не все…
Приведу вам пример.
Допустим что какой-то человек пришёл в спортзал. Причём до этого он никогда и ничем не занимался и поэтому на данный момент его максимальная мышечная активность находиться на уровне 30% – 40% процентов от их максимального 100% процентного потенциала на данный момент тех мышц, которые он имеет сейчас.

- Зелёная полоса — показывает мышечную активность на уровне 5% — 10% процентов применяемую в повседневной жизни.
- Синяя полоса – показывает мышечную активность (силу) не тренированного человека на уровне 30% — 40% процентов.
- Красная полоса — показывает максимальный 100% процентный потенциал мышц и их возможность на данный момент.
- Голубая полоса — показывает прохождение вашего силового максимума и последующую гипертрофию ваших мышц.
И если он не увеличит силу своих мышц и не вовлечёт мышечную активность на все 100% процентов, тогда никакой гипертрофии в дальнейшем просто не будет.

Если же он будет увеличивать силу своих мышц, то и мышечная активность будет тоже расти до отметки в 100% процентов и как только он дойдёт до этой отметки и превысит её, то только после этого последует гипертрофия мышц, то есть начнётся увеличение самих мышц.
Давайте попытаемся с вами разобраться, почему на начальном этапе своих тренировок всегда следует работать только лишь на увеличение своих силовых показателей.
Вернёмся к тому, что человек только недавно начал заниматься и его максимальный жим лёжа на данный момент составляет 50 кг, но его мышцы способны и могут выжать 80 кг, но на данный момент он этого сделать не может, сейчас он может выполнить только лишь 50 кг в своём жиме лёжа.
Получается что на данный момент эти самые 80 кг являются по сути его мышечным потенциалом. Иными словами эти 80 кг это и есть его 100% мышечный потенциал и та мышечная активность с которой его мышцы способны справиться в этом упражнение.

И условно говоря, пока он не дойдёт до этих 80 кг в жиме лёжа его мышцы не будут расти потому как он не исчерпал свой мышечный потенциал, при котором его мышцы способны работать на данный момент, не прибегая при этом к гипертрофии и увеличению самого объёма его мышечных волокон.
И лишь тогда когда он дойдёт до этого веса и преодолеет его, только лишь после этого начнётся сам процесс мышечного роста потому как произойдёт перегруз его мышечной активности за 100% процентов.
- Именно поэтому все новички приходя в спортзал способны за несколько месяцев прибавить сразу 30 – 40 кг и даже более, к примеру, в том же в жиме лёжа, а также в других упражнениях.
- И именно поэтому в самом начале все начинающие атлеты так стремительно прибавляют в течение целого года свои силовые показатели, увеличивая их чуть ли не вдвое.
Это связано с тем, что до спортзала они уже имели изначально не выработанный потенциал своих мышц, которые они уже включили лишь тогда, когда начали активно заниматься и увеличивать свои силовые показатели и увеличивать силу своих мышц.
Но я огорчу вас, потому что этот период стремительного роста ваших силовых это всего лишь временный период, за которым уже следует тот предел и та планка, которую многие потом очень долгое время не могут преодолеть.
Именно поэтому многие новички, достигнув определённого роста своих силовых возможностей в том или ином упражнении, потом очень долгое время не могут больше прибавить в своих силовых.
А всё потому, что до этого занимаясь даже абы как, вы всё равно прибавляли и увеличивали свои силовые показатели, но затем как только вы дошли до определённого предела, т.е. до той точки 100% процентного мышечного потенциала, который вы изначально кстати уже имели, то после этого вы долгое время уже не можете потом его преодолеть.
Причём это может доходить и до нескольких лет пустых тренировок, когда атлет не может прибавить, к примеру, в своём жиме лёжа даже пару кг и с этой проблемой наверняка сталкивались очень многие из вас.
Для того чтобы всего этого избежать, в первую очередь вам нужно научиться грамотно строить свои тренировки и свои тренировочные циклы и только тогда вы будете увеличивать и дальше свои силовые показатели и сможете всегда прогрессировать в мышечной массе.
Энергетика мышечной деятельности.
Одно мышечное волокно может содержать 15 миллиардов толстых нитей. При том, что мышечные волокна активно сокращаются, в каждой толстой нити распадаются примерно 2500 молекул АТФ (нуклеотид, играющий важную роль в обмене энергии и веществ в организме) в секунду. Даже небольшие скелетные мышцы содержат тысячи мышечных волокон.
Основной функцией АТФ является передача энергии из одного места в другое, а не долгосрочное хранение энергии. В состоянии покоя скелетные мышечные волокна производят больше АТФ, чем они нуждается. В этих условиях АТФ передает энергию креатина. Креатин представляет собой небольшую молекулу, которую мышечные клетки собирают из фрагментов аминокислот. Передача энергии создает еще одно высокоэнергетическое соединение креатин фосфата (КФ).
АТФ + креатин АДФ + креатин фосфат
Во время мышечного сокращения происходит разрыв соединений АТФ, в результате этого образуется аденозиндифосфат (АДФ). Энергия, запасенная в креатинфосфате затем используется для «перезарядки» АДФ, превращая его обратно в АТФ через обратную реакцию.

АДФ + креатин фосфат + креатин
Фермент креатинфосфокиназа (КФК) облегчает эту реакцию. Когда мышечные клетки повреждены, происходит утечка КФК через клеточные мембраны в кровоток. Таким образом, высокая концентрация в крови КФК обычно указывает на серьезное повреждение мышц.
Отдыхающие скелетные мышечные волокна содержат примерно в шесть раз больше креатин фосфата как АТФ. Но когда мышечные волокна переживают устойчивое напряжение, эти энергетические запасы будут исчерпаны всего лишь примерно за 15 секунд. Мышечные волокна должны затем полагаться на другие механизмы для преобразования АДФ в АТФ.
Большинство клеток в организме генерируют АТФ через аэробный метаболизм в митохондриях и через гликолиз в цитоплазме. Аэробный метаболизм (сопровождающийся потреблением кислорода) обычно обеспечивает 95 % АТФ в покоящейся клетке. В этом процессе митохондрии поглощают кислород, АДФ, ионы фосфата и органические субстраты от окружающей цитоплазмы. Субстраты затем вводят цикл трикарбоновых кислот (также известный как цикл лимонной кислоты или цикл Кребса), ферментативный путь, который разрушает органические молекулы. Атомы углерода выпускаются, как двуокись углерода, атомы водорода курсируются дыхательными ферментами во внутренней митохондриальной мембране, где их электроны удаляются. После серии промежуточных шагов, протоны и электроны сочетаются с кислородом и образованием воды. В этом эффективном процессе высвобождается большое количество энергии и используется для создания АТФ.

Отдыхающие скелетные мышечные волокна полагаются почти исключительно на аэробный метаболизм жирных кислот, чтобы генерировать АТФ. Когда мышца начинает сжиматься, митохондрии начинают разрушение молекулы пировиноградной кислоты вместо жирных кислот. Пировиноградная кислота обеспечивается ферментативным путем гликолиза. Гликолиз является распадом глюкозы до пировиноградной кислоты в цитоплазме клетки. Этот процесс называется анаэробным, потому что он не требует кислорода. Гликолиз обеспечивает увеличение АТФ и генерирует 2 молекулы пировиноградной кислоты из каждой молекулы глюкозы. АТФ образуется в процессе гликолиза. Поскольку гликолиз может протекать в отсутствие кислорода, он может быть особенно важен, когда наличие кислорода ограничивает скорость производства митохондриальной АТФ. В большинстве скелетных мышц гликолиз является основным источником АТФ во время пиковых периодов активности. Расщепление глюкозы в этих условиях происходит в основном из резервов гликогена в саркоплазме. Гликоген представляет собой полисахарид цепочек молекул глюкозы. Типичные волокна скелетных мышц содержат большие запасы гликогена, которые могут составлять 1,5 % от общего веса мышц.

Энергопотребление и уровень мышечной активности.
В скелетных мышцах, при нахождении их в состоянии покоя, спрос на АТФ является низким. Более чем достаточно доступного кислорода митохондриям для удовлетворения этого спроса, в итоге они производят избыток АТФ. Дополнительный АТФ используется для создания запасов гликогена. Отдыхающие мышечные волокна поглощают жирные кислоты и глюкозу, которые доставляются кровотоком. Жирные кислоты расщепляются в митохондриях и АТФ генерируется для преобразования креатина в креатин фосфат и глюкозы в гликоген.
При умеренных уровнях физической активности увеличивается потребность в АТФ. Этот спрос удовлетворяется за счет митохондрий, когда скорость производства митохондриальной АТФ повышается, что увеличивает скорость потребления кислорода. Наличие кислорода не является ограничивающим фактором, потому что кислород может диффундировать (соединяться, смешиваться) в мышечном волокне достаточно быстро, чтобы удовлетворить митохондриальные потребности. Скелетные мышцы в этот момент зависят главным образом от аэробного метаболизма пировиноградной кислоты, чтобы генерировать АТФ. Пировиноградная кислота образуется в процессе гликолиза, который расщепляет молекулы глюкозы, полученные из гликогена в мышечных волокнах. Если запасы гликогена низки, мышечное волокно может также расщипить и другие субстраты, такие как липиды или аминокислоты. Пока спрос на АТФ может быть удовлетворен путем митохондриальной активности, обеспечивание АТФ гликолизом остается незначительным в общем энергетическом процессе мышечного волокна.

В пиковые уровни активности требуется много АТФ, в результате чего производство АТФ в митохондриях возрастает до максимума. Это максимальная скорость определяется наличием кислорода, а кислород не может диффундировать в мышечных волоконах достаточно быстро, чтобы дать возможность митохондрии для образования требуемого АТФ. В пиковых уровнях нагрузки митохондриальная активность может обеспечить лишь около одной трети от необходимого АТФ. Остальная часть приходится на гликолиз.
Когда гликолиз производит пировиноградную кислоту быстрее, чем она может быть использована в митохондрии, увеличивается уровень пировиноградной кислоты в саркоплазме. В этих условиях пировиноградная кислота превращается в молочную кислоту.
Анаэробный процесс гликолиза позволяет клетке генерировать дополнительный АТФ, когда митохондрии не в состоянии удовлетворить текущие потребности в энергии. Тем не менее, производство анаэробной энергии имеет свои недостатки:
Молочная кислота представляет собой органическую кислоту, которая в жидкостях организма
диссоциирует на ионы водорода и отрицательно заряженного иона лактата. Таким образом, производство молочной кислоты может привести к снижению внутриклеточного рН. Буферы в саркоплазме могут противостоять рН сдвигам, но эти защиты ограничены. В конце концов изменения рН будут изменять функциональные характеристики ключевых ферментов.
Гликолиз является относительно неэффективным способом для генерации АТФ. В анаэробных условиях каждая молекула глюкозы генерирует 2 молекулы пировиноградной кислоты, которые превращаются в молочную кислоту. В свою очередь, клетка получает 2 молекулы АТФ через гликолиз. Если бы те молекулы пировиноградной кислоты катаболизировались бы аэробным способом в митохондриях, клетка получила бы 34 дополнительных молекул АТФ.

Мышечная усталость.
Скелетные мышечные волокна устают тогда, когда они больше не могут сокращаться, несмотря на продолжение нервного импульса. Причина мышечной усталости изменяется в зависимости от уровня активности мышц. После коротких пиковых уровней деятельности, например таких, как 100-метровый забег на время, усталость может быть
результатом исчерпания запасов АТФ или от падения рН, который сопровождается накоплением молочной кислоты. После длительных напряжений, таких как марафон, усталость может включать физические повреждения саркоплазматического ретикулума, что мешает регуляции внутриклеточных + концентрации ионов Ca2. Мышечная усталость накапливается и последствия этого становятся более выраженными, поскольку все больше мышечных волокон начинают задействоваться этим состоянием. Результатом является постепенное снижение возможностей всех скелетных мышц.

Если мышечное волокно сокращается при умеренных уровнях и запросы АТФ могут быть удовлетворены через аэробный метаболизм, усталость не произойдет, пока запасы гликогена, липидов и аминокислот не будут исчерпаны. Этот тип усталости происходит с мышцами спортсменов долгих нагрузок, таких как марафонцы, после нескольких часов забегов на длинные дистанции.
Когда мышца производит внезапный, интенсивный всплеск активности на пиковых уровнях, большая часть АТФ обеспечивается путем гликолиза. После нескольких секунд до минуты, повышение уровня молочной кислоты снижает рН тканей и мышцы больше не могут функционировать нормально. Спортсмены, которые испытывают быстрые мощные нагрузки, такие как спринтеры в 100-метровом забеге, получают как раз этот тип мышечной усталости.

Для нормальной функциональности мышц требуется:
1) существенные внутриклеточные энергетические запасы, 2) нормальное кровообращение и 3) нормальная концентрация кислорода в крови. Все, что препятствует одному или более из этих факторов, будет способствовать преждевременной усталости мышц. Например, снижение кровотока от тесной одежды, расстройство кровообращения или потеря крови замедляет доставку кислорода и питательных веществ, ускоряя при этом накопление молочной кислоты, а также способствует мышечной усталости.
Восстановительный период. При сокращении мышечных волокон условия в саркоплазме меняются. Потребляются энергетические запасы, выделяется тепло и, если сокращение было пиковым, генерируется молочная. В период восстановления, условия в мышечных волокнах возвращаются к нормальным. Может занять несколько часов для того,чтобы мышечные волокона оправились от периода умеренной активности. После длительной деятельности на более высоких уровнях активности, полное восстановление может занять неделю. В период восстановления, когда кислорода имеется в изобилии, молочная кислота может быть переработана путем конвертации обратно в пировиноградную кислоту.
Пировиноградная кислота может быть использована или митохондрией для генерации АТФ, или в качестве субстрата для ферментов, которые синтезируют глюкозу и восстанавливают запасы гликогена.

В период нагрузок молочная кислота диффундирует из мышечных волокон в кровоток. Этот процесс продолжается после того, как напряжение закончилось, потому что внутриклеточные концентрации молочной кислоты все еще относительно высоки. Печень поглощает молочную кислоту и преобразует ее в пировиноградную кислоту. Приблизительно 30% этих молекул пировиноградной кислоты расщепляются, обеспечивая АТФ, необходимый для превращения других молекул пировиноградной кислоты в глюкозу. Молекулы глюкозы затем выпускаются в обращение, где они поглощаются скелетными мышечными волокнами и используются для восстановления их запасов гликогена. Эта перетасовка молочной кислоты в печени и глюкозы к мышечным клеткам называется циклом Кори.
В период восстановления кислород легко доступен и потребность тела в кислороде остается повышенной, выше нормального уровня покоя. Восстановительный период подпитывается от АТФ. Чем больше АТФ требуется, тем больше кислорода будет необходимо. Кислородный долг или избыточное послетренировочное потребление кислорода, созданное во время физических упражнений, является тем самым количеством кислорода, которое необходимо для нормального восстановления. Скелетные мышечные волокна, которые должны восстановить АТФ, креатинфосфат и гликоген, в концентрации своих прежних уровнях и клетки печени, которые генерируют АТФ, необходимый для превращения избыточной молочной кислоты в глюкозу, несут ответственность за большую часть дополнительного потребления кислорода. В то время, как кислородный долг восполняется, частота и глубина дыхания увеличиваются. В результате, вы будете продолжать дышать тяжело достаточно долго после прекращения интенсивных тренировок.

Тепловые потери мышечной активности генерирует значительные объемы тепла. Когда происходит катаболическая реакция, например, при пробое гликогена или реакций гликолиза, мышечные волокна захватывают только часть выделенной энергии. Остальная высвобождается в виде тепла. Отдыхающие мышечные волокна, опирающиеся на аэробный метаболизм, захватывают около 42 % энергии, выделяемой в катаболизме. Другие 58 % согревают саркоплазму тканевой жидкости и циркулирующую кровь. Активные скелетные мышцы выпускают около 85 % тепла, необходимого для поддержания нормальной температуры тела.
Когда мышцы становятся активными, их энерго-потребление резко возрастает. Поскольку производство анаэробной энергии становится основным методом АТФ, мышечные волокна уже менее эффективно поглощают энергию. В пиковых уровнях нагрузки, только около 30 % от выделенной энергии сохраняется в виде АТФ, а остальные 70 % согревают мышцы и окружающие их ткани.
Гормоны и мышечный метаболизм. Метаболическая активность в скелетных мышечных волокнах регулируется гормонами эндокринной системы. Гормон роста из гипофиза и тестостерон (основной половой гормон у мужчин) стимулируют синтез сократительных белков и расширение скелетных мышц. Гормоны щитовидной железы поднимают скорость потребления энергии при отдыхе. Во время интенсивной физической активности, гормоны надпочечники, особенно адреналин, стимулируют мышечный обмен веществ и увеличивают продолжительность стимуляции и силу сокращения.
В. Н. Селуянов, В. А. Рыбаков, М. П. Шестаков
Глава 1. Модели систем организма
1.1.4. Физиология мышечной деятельности
Биохимия и физиология мышечной активности при выполнении физической работы может быть описана следующим образом. Покажем с помощью имитационного моделирования как разворачиваются физиологические процессы в мышце при выполнении ступенчатого теста.
Предположим, что мышца (например, четырехглавая мышца бедра) имеет ММВ 50 %, амплитуда ступеньки - 5 % максимальной алактатной мощности, величина которой принята за 100 %, длительность - 1 мин. На первой ступеньке в связи с малым внешним сопротивлением рекрутируются, согласно «правилу размера» Ханнемана, низкопороговые ДЕ (МВ). Они имеют высокие окислительные возможности, субстратом в них являются жирные кислоты. Однако первые 10 20 с энергообеспечение идет за счет запасов АТФ и КрФ в активных МВ. Уже в пределах одной ступеньки (1 мин.) имеет место рекрутирование новых мышечных волокон, благодаря этому удается поддерживать заданную мощность на ступеньке. Вызвано это снижением концентрации фосфогенов в активных МВ, то есть силы (мощности) сокращения этих МВ, усилением активирующего влияния ЦНС, а это приводит к вовлечению новых ДЕ (МВ). Постепенное ступенчатое увеличение внешней нагрузки (мощности) сопровождается пропорциональным изменением некоторых показателей: растет ЧСС, потребление кислорода, легочная вентиляция, не изменяется концентрация молочной кислоты и ионов водорода.
При достижении внешней мощности некоторого значения наступает момент, когда в работу вовлекаются все ММВ и начинают рекрутироваться промежуточные мышечные волокна (ПМВ). Промежуточными мышечными волокнами можно назвать те, в которых массы митохондрий недостаточно для обеспечения баланса между образованием пирувата и его окислением в митохондриях. В ПМВ после снижения концентрации фосфогенов активизируется гликолиз, часть пирувата начинает преобразовываться в молочную кислоту (точнее говоря, в лактат и ионы водорода), которая выходит в кровь, проникает в ММВ. Попадание в ММВ (ОМВ) лактата ведет к ингибированию окисления жиров, субстратом окисления становится в большей мере гликоген. Следовательно, признаком рекрутирования всех ММВ (ОМВ) является увеличение в крови концентрации лактата и усиление легочной вентиляции. Легочная вентиляция усиливается, в связи с образованием и накоплением в ПМВ ионов водорода, которые при выходе в кровь взаимодействуют с буферными системами крови и вызывают образование избыточного (неметаболического) углекислого газа. Повышение концентрации углекислого газа в крови приводит к активизации дыхания (Физиология человека, 1998).
Таким образом, при выполнении ступенчатого теста имеет место явление, которое принято называть аэробным порогом (АэП). Появление АэП свидетельствует о рекру-тировании всех ОМВ. По величине внешнего сопротивления можно судить о силе ОМВ, которую они могут проявить при ресинтезе АТФ и КрФ за счет окислительного фосфори-лирования (Селуянов В. Н. с соав., 1991).
Дальнейшее увеличение мощности требует рекрутирования более высокопороговых ДЕ (ГМВ), в которых митохондрий очень мало. Это усиливает процессы анаэробного гликолиза, больше выходит лактата и ионов Н в кровь. При попадании лактата в ОМВ он превращается обратно в пируват с помощью фермента ЛДГ Н (Karlsson, 1971,1982). Однако мощность митохондриальной системы ОМВ имеет предел. Поэтому сначала наступает предельное динамическое равновесие между образованием лактата и его потреблением в ОМВ и ПМВ, а затем равновесие нарушается, и некомпенсируемые метаболиты - лактат, Н, СО 2 - вызывают резкую интенсификацию физиологических функций. Дыхание один из наиболее чувствительных процессов, реагирует очень активно. Кровь при прохождении легких в зависимости от фаз дыхательного цикла должна иметь разное парциальное напряжение СО 2 . «Порция» артериальной крови с повышенным содержанием СО 2 достигает хеморецепторов и непосредственно модулярных хемочувствительных структур ЦНС, что и вызывает интенсификацию дыхания. В итоге СО 2 начинает вымываться из крови так, что в результате средняя концентрация углекислого газа в крови начинает снижаться. При достижении мощности, соответствующей АнП, скорость выхода лактата из работающих гликолитических МВ сравнивается со скоростью его окисления в ОМВ. В этот момент субстратом окисления в ОМВ становятся только углеводы (лактат ингибирует окисление жиров), часть из них составляет гликоген ММВ, другую часть - лактат, образовавшийся в гликолитических МВ. Использование углеводов в качестве субстратов окисления обеспечивает максимальную скорость образования энергии (АТФ) в митохондриях ОМВ. Следовательно, потребление кислорода или (и) мощность на анаэробном пороге (АнП) характеризует максимальный окислительный потенциал (мощность) ОМВ (Селуянов В. Н. с соав., 1991).
Дальнейший рост внешней мощности делает необходимым вовлечение все более высокопороговых ДЕ, иннервирующих гликолитические МВ. Динамическое равновесие нарушается, продукция Н, лактата начинает превышать скорость их устранения. Это сопровождается дальнейшим увеличением легочной вентиляции, ЧСС и потребления кислорода. После АнП потребление кислорода в основном связано с работой дыхательных мышц и миокарда. При достижении предельных величин легочной вентиляции и ЧСС или при локальном утомлении мышц потребление кислорода стабилизируется, а затем начинает уменьшаться. В этот момент фиксируют МПК.